 |
| |||
Med Sci (Paris). 2014 November; 30(11): 980–990. Published online 2014 November 10. doi: 10.1051/medsci/20143011012.Le cil primaire, orchestrateur de la morphogenèse cérébrale 1Laboratoire de biologie du développement, UPMC Université Paris 6, UMR 7622 CNRS, U969 Inserm, 9, quai Saint Bernard, 75005Paris, France Corresponding author. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||
Le cerveau des mammifères est un organe hautement sophistiqué dont la morphologie reflète la diversité régionale des réseaux cellulaires. Cette fascinante complexité a été à l’origine de très nombreuses études visant à mieux comprendre les différentes étapes de la construction cérébrale. Cependant, ce n’est que récemment que le rôle du cil primaire a été mis à jour. Certes, ces petites extensions cellulaires structurées par un axonème (9 doublets de microtubules) et un corps basal (centriole père du centrosome) [ 65, 66] (→) avaient été décrites dans le cerveau des vertébrés dans les années 1960 grâce à l’avènement de la microscopie électronique à transmission. Mais, à l’époque, les cils étaient considérés comme des organites vestigiaux sans fonction particulière [ 1– 3]. Ce n’est qu’avec l’analyse des lignées de souris dépourvues de cil et la description des syndromes humains dus à des dysfonctionnements ciliaires, que les fonctions de ce petit organite solitaire se sont révélées essentielles à de nombreux processus développementaux et, notamment, à la morphogenèse cérébrale [ 4, 5]. (→) Voir les Synthèses de C. Fort et P. Bastin ; de A. Benmerah, pages 955 et 962 de ce numéro Les ciliopathies sont des maladies génétiques dues à des dysfonctionnements du cil d’origine variable [ 6, 67] (→). Sur la base de phénotypes communs, on y associe des syndromes résultant d’anomalies des voies de signalisation en aval du cil [ 7]. De nombreuses anomalies du système nerveux central (SNC) et de nombreuses manifestations cliniques neurologiques ont été décrites dans les ciliopathies (Tableau I) [ 8]. Cette variabilité s’explique par la nature de la mutation et/ou les atteintes combinées de plusieurs gènes. Certaines caractéristiques neuroradiologiques font partie des critères diagnostiques, comme le signe de « la dent molaire » (molar tooth sign, voir Tableau I ) chez les patients atteints du syndrome de Joubert ou apparenté (JSRD, Joubert syndrome related disorders) [67] (→). (→) Voir la Synthèse de R. Bachmann-Gagescu, page 1011 de ce numéro
Certaines atteintes peuvent être fréquentes, sans être constantes ; c’est le cas des agénésies du corps calleux dans les syndromes OFD (oral-facial-digital), acrocalleux et hydroléthalus ou, plus rarement, dans le syndrome de Joubert. Très fréquemment, des troubles neurologiques, cognitifs ou comportementaux ont été décrits, sans pour autant que les bases neuro-anatomiques aient été systématiquement analysées. Dans tous les cas, une origine neuro-développementale de ces atteintes est possible, mais reste à déterminer. Comprendre comment se forme le cerveau au cours du développement embryonnaire est indispensable à la compréhension de son fonctionnement et, le cas échéant, de ses dysfonctionnements. Parmi les nombreuses approches possibles, l’analyse des phénotypes mutants, chez des modèles murins en particulier, est celle qui a le plus apporté à l’étude de la morphogenèse cérébrale [ 9]. Rappelons que les mutants ciliaires, par définition, présentent une altération de la structure et/ou de la fonction du cil, qui se manifeste chez la souris par une association de phénotypes récurrents : défauts de latéralité et défauts de régionalisation (patterning) des bourgeons de membres et du tube neural spinal [ 10]. Cette triade constitue la signature phénotypique des mutants ciliaires, et les autres phénotypes, en particuliers cérébraux, ont parfois été occultés. Surtout, nombre de ces mutants ciliaires meurent en milieu de gestation (entre E10 et E13 chez la souris). Dans ce cas, l’analyse des étapes ultérieures du développement ne peut se faire qu’en analysant des mutants hypomorphes ou des animaux chez lesquels un knock-out (KO) conditionnel du gène est réalisé. Cette revue fait le point sur les données, les hypothèses et les questions relatives au rôle du cil primaire dans la formation du cerveau et, plus particulièrement, du cerveau antérieur. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
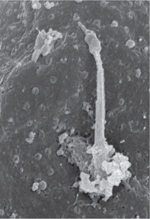
Les milliards de neurones et de cellules gliales qui constituent le cerveau des mammifères sont générés à partir du feuillet ectodermique de l’embryon. La plaque neurale, induite lors de la gastrulation, contient les cellules neuro-épithéliales, dont les changements de forme et d’adhésivité vont contribuer à la formation du tube neural. Dès le début de la neurulation (E8,0 chez la souris), ces cellules arborent un cil primaire sur leur face apicale (Figure 1A). Ces cils baignent initialement dans la cavité amniotique, puis feront face ensuite à la cavité centrale du tube neural après le soulèvement et la fusion des bourrelets neuraux le long de la ligne médiane.
Cil primaire et fermeture du tube neural Si la fermeture du tube neural ne s’opère pas correctement, le développement ultérieur du système nerveux central s’en trouve compromis. Or, plusieurs mutants ciliaires présentent un phénotype attestant d’un défaut de fermeture du tube neural antérieur (exencéphalie), qui s’accompagne d’un développement extériorisé du cerveau chez la souris : Dynchc2/Dync2h1 (dynein, cytoplasmic 2, heavy chain 1), Ift52 (intraflagellar transport protein 52), Ift57hippi, Ift88flexo, Ift122, Ift172wimple, Mks1 (Meckel syndrome, type 1), Inpp5e (inositol polyphosphate-5-phosphatase), Ftm/Rpgrip1l (retinitis pigmentosa GTPase regulator interacting protein 1l), Tmem67 (transmembrane protein 67), Tulp3 (Tubby-like protein 3), etc. [
11]. Chez l’homme, des défauts équivalents conduisent à des anencéphalies, un signe clinique rare, décrit parfois chez des fœtus atteints du syndrome de Meckel ou du syndrome acrocalleux [
2]. Ainsi, le cil primaire semble impliqué dans la fermeture du tube neural antérieur de la souris, mais par quels mécanismes ? Cela reste à établir précisément [
12]. Il est à noter également que ces mutations ne sont pas totalement pénétrantes. L’incidence de l’exencéphalie est, par exemple, de 77 % chez les souris Tulp3-/-
, 37 % pour la mutation Ftm-/-, 30 % pour la mutation Inpp5e-/-, ou encore 28 % chez les animaux Mks1-/-
[11]. De plus, le fond génétique influence considérablement la fréquence de ce phénotype, ce qui pourrait indiquer un déterminisme multifactoriel et/ou des sensibilités différentes aux subtiles variations stochastiques qui surviennent lors du développement.Dès sa formation, le tube neural présente des différences morphologiques le long des axes antéro-postérieur (AP) et dorso-ventral (DV). La partie antérieure du tube neural se caractérise par des renflements clairement visibles, qui correspondent aux futures subdivisions morphologiques du cerveau (Figure 1B). La partie postérieure, plus fine, est à l’origine de la moelle épinière. Les processus de régionalisation sont conditionnés par des signaux morphogènes et l’expression de gènes régulateurs (facteurs de transcription) qui dictent le destin et le comportement ultérieur des cellules neurales. Le cil primaire, un acteur clé de la signalisation Hedgehog Dans le tube neural postérieur, la spécification des types cellulaires ventraux est dépendante de la production de sonic hedgehog (Shh, principale protéine de la famille Hh produite dans le SNC). Shh est sécrétée par les cellules ventrales de la plaque du plancher et constitue un gradient morphogène ventralisant [
13]. Or, chez un grand nombre de mutants ciliaires, une réduction, voire une absence, de spécification de certains types cellulaires ventraux ont été décrites [
68] (→).(→) Voir la Synthèse de C. Métin, page 991 de ce numéro Cette dorsalisation du tube neural spinal atteste d’un défaut d’activation de la voie de signalisation Hh dans les territoires ventraux [ 14]. C’est dans ce contexte développemental que le cil primaire s’est révélé être un élément essentiel dans la perception et la transduction du signal Hh chez les mammifères [14, 15]. Le cil est alors apparu comme une « antenne cellulaire » dédiée à la réception des signaux de l’environnement. Cependant, en absence de signal Hh, le cil primaire contrôle l’activité des effecteurs transcriptionnels de la voie Hh que sont les protéines Gli (famille de facteurs de transcription à doigt de zinc associées aux gliomes, voir Figure 2 ). En particulier, le clivage protéolytique de la protéine Gli3, qui conditionne son activité répresseur, est dépendant de l’intégrité ciliaire. Ainsi, une réduction de la forme tronquée de la protéine Gli3 (Gli3-R) est fréquemment observée chez les mutants ciliaires. Cette fonction du cil, indépendante de Hh, est déterminante pour la régionalisation dorso-ventrale dans la partie antérieure du tube neural et antéro-postérieure dans les bourgeons des membres. En effet, plusieurs mutants ciliaires présentent des défauts de régionalisation dorso-ventrale du télencéphale. Il s’agit de mutants déficients pour différents gènes, tous impliqués dans la formation ou la fonction du cil, codant pour des protéines apposées au corps basal (Ftm/Rpgrip1l, Ofd1), des IFT (Ift88Cobblestone, Ift139alien, Ift172) ou des sous-unités de moteurs moléculaires (Dynchc2, Kif3a [kinesin family member 3A]) impliqués dans le transport intraflagellaire antérograde (Ift88, Ift172, Kif3a) ou rétrograde (Ift139, Dynchc2) [ 16– 21, 65] (→). (→) voir la synthèse de C. Fort et P. Bastin, page 955 de ce numéro
Tous ces mutants présentent une ventralisation du télencéphale, c’est-à-dire une extension du territoire ventral (subpallium) au détriment du territoire dorsal (pallium), et la frontière entre les deux s’estompe (présence ectopique de cellules d’identité ventrale dans le territoire dorsal) (Figure 1). Ainsi, l’altération du cil peut conduire à la fois à une ventralisation du télencéphale et à une dorsalisation du tube neural postérieur. Ceci s’explique par le fait que les effecteurs principaux de la voie Hh ne sont pas les mêmes dans ces deux régions du SNC (rôle majeur de Gli2 dans le tube neural spinal et de Gli3 dans le télencéphale). La ventralisation du télencéphale des mutants ciliaires ressemble à ce qui a été décrit chez les mutants Gli3 [ 22], anomalies qui avaient conduit à définir cet effecteur de la voie Hh comme un acteur majeur de la spécification du télencéphale dorsal [9]. Ainsi, il semblait raisonnable de postuler que les cils primaires étaient nécessaires à la spécification du télencéphale dorsal via leur rôle dans la production de Gli3-R. Nous avons formellement validé cette hypothèse en réintroduisant la protéine Gli3-R chez les mutants ciliaires Rpgrip1lFtm/Ftm en croisant ces souris avec la lignée Gli3∆699 qui exprime une forme tronquée de Gli3 fonctionnellement semblable à Gli3-R (knock-in Gli3∆699 au locus Gli3) [23]. L’introduction d’un allèle Gli3∆699 chez les mutants Rpgrip1lFtm/Ftm est suffisante pour rétablir la régionalisation dorso-ventrale du télencéphale à E12,5. Cette correction phénotypique a lieu malgré la persistance des altérations ciliaires et montre qu’il est possible de restaurer la fonction des cils primaires in vivo en ciblant l’effecteur maître en aval des cils, ici Gli3-R. De plus, par la même approche, nous avons montré que la morphogenèse des bulbes olfactifs, altérée par la ventralisation du télencéphale chez le mutant Rpgrip1lFtm/Ftm , est restaurée par l’apport de Gli3-R (par l’allèle Gli3∆699 ) [20]. Dans un autre contexte, Gli3∆699 corrige le phénotype rénal des mutants Kif3a [ 24]. Cependant, Gli3∆699 ne corrige pas tous les phénotypes des mutants ciliaires et, au contraire, pourrait exacerber certains défauts (au niveau des membres par exemple). Une autre tentative de correction est illustrée par l’étude du mutant Ift139alien . Le phénotype de ce dernier présente la particularité, au niveau du tube neural postérieur, d’être corrélé à une activation accrue de la voie Hh (ventralisation), et non pas à une réduction comme c’est le cas chez la plupart des autres mutants [ 25]. Ici, le but a donc été de réduire la signalisation Hh, en croisant les souris Ift139alien avec les lignées KO pour Shh ou Gli2. Les corrections phénotypiques observées chez les doubles mutants Ift139alien/alien Shh+/- et Ift139alien/alien Gli2+/- , respectivement dans le télencéphale et le tube neural postérieur, semblent conforter l’idée que le défaut résulte ici d’un excès de signal Hh et d’une surproduction de la forme activatrice de Gli2 [ 19, 25]. Des défauts ciliaires pourraient ainsi conduire, selon le type de mutation, soit à l’activation de l’activateur Gli2 (Ift139alien ), soit à l’inhibition du répresseur Gli3 (Ftm/Rpgrip1l). Or, l’équilibre entre ces deux formes actives des protéines Gli (activateur/répresseur) est déterminant dans de nombreux processus développementaux. D’autres défauts cérébraux précoces ont été décrits chez les mutants ciliaires, comme une holoprosencéphalie semilobaire (défaut de septation médiane du télencéphale) chez les mutants Ift172 et Ttbk2 (définie au stade E10,5) [ 17, 26]. De plus, des défauts de frontière le long de l’axe antéro-postérieur ont été rapportés : entre le télencéphale et le diencéphale chez les mutants Ift88cobblestone et Ift139alien [ 18, 19], et entre le mésencéphale et le métencéphale dans le knock-out Ift172 [17]. Ces défauts de régionalisation peuvent avoir des conséquences dramatiques sur les étapes suivantes de la construction du cerveau embryonnaire. En effet, les différentes régions spécifiées possèdent d’emblée des propriétés spécifiques qui conditionnent leur développement ultérieur en termes de prolifération, différenciation, migration cellulaire, navigation axonale et synaptogenèse. Le cil primaire semble intervenir également dans le contrôle de ces différents comportements cellulaires, indépendamment de son rôle dans la régionalisation. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
Depuis la neurulation et jusqu’à la naissance, quasiment toutes les cellules du SNC en développement arborent un cil primaire : pas seulement les neurones différenciés, mais également les progéniteurs neuraux en phase de prolifération et les précurseurs d’interneurones pendant leur migration (Figure 3).
Cil primaire et transition prolifération/différenciation des progéniteurs neuraux Dans les progéniteurs neuraux, le centrosome est alternativement requis pour ancrer le cil primaire (pendant l’interphase) et pour organiser les fuseaux de division (en mitose) ; le corps basal du cil n’est autre que le centriole père du centrosome. Il existe donc un lien étroit entre le cil et le cycle cellulaire [
27,
69] (→).(→) Voir la Synthèse de N. Delgehyr et N. Spassky, page 976 de ce numéro Cependant, selon les modèles cellulaires considérés, les conclusions semblent diverger. Dans certains cas, la présence du cil ralentirait la progression du cycle, ce dernier s’accélérant chez les mutants ciliaires. C’est ce qui a été montré dans le cortex du mutant conditionnel nestin-Cre ; Kif3aflox [21]. Dans d’autres cas, la présence du cil est nécessaire pour maintenir un nombre important de cellules en division, via la transduction du signal Shh qui a un effet mitogène dans ce contexte. C’est le cas, par exemple, des progéniteurs des grains dans le cervelet, dont le nombre est réduit chez les mutants conditionnels Ift88flox et Kif3flox (dont l’invalidation est ciblée par hGFAP [human glial fibrillary acidic protein]-Cre) [ 28, 29]. Ces défauts sont à corréler avec les hypoplasies du vermis cérébelleux (région centrale du cervelet) décrites dans plusieurs ciliopathies (BBS [Bardet-Biedl syndrome], OFD, néphronophtise, syndrome de Jeune) [ 30] (Tableau I ). En plus de son influence directe sur la durée des cycles cellulaires et sur le nombre de cellules en division, le cil primaire serait un élément essentiel dans l’équilibre entre divisions symétriques/asymétriques lors de la neurogenèse corticale (Figure 3). Lors des divisions asymétriques, la cellule fille, qui hérite du centriole père le plus âgé, reforme plus rapidement un cil primaire, qui d’ailleurs ne disparaît pas forcément totalement pendant la mitose [ 31]. Elle pourrait ainsi répondre plus rapidement aux signaux tels que Shh [ 32]. De plus, le centriole père, le plus âgé, ségrège plus fréquemment dans les cellules gardant leur caractère progéniteur (plutôt que dans les cellules s’engageant dans la différenciation neuronale) [ 33]. En conclusion, les cils primaires, par leur interaction avec le centriole père du centrosome, sont liés à la transition prolifération/différenciation des progéniteurs neuraux qui influencent directement la neurogenèse corticale. Le cil primaire, producteur de signaux ? Dès le début de la neurogenèse, les progéniteurs neuraux, aussi appelés cellules neuroépithéliales, forment un épithélium pseudostratifié homogène : toutes les cellules présentent les mêmes caractéristiques de polarité apicobasale, le cil se trouvant sur la face apicale. Les futurs neurones corticaux se détachent ensuite de la zone apicale pour migrer et se différencier en périphérie. Un des tout premiers signes distinctifs de l’engagement des cellules vers un destin neuronal serait la formation d’un cil en position basolatérale, et non plus en position apicale [
34]. Récemment, une étude réalisée au niveau du tube neural postérieur a montré l’existence d’un mécanisme d’abscission apicale, jusque là inconnu [
35]. La partie apicale de la cellule, comprenant le cil et les jonctions serrées, reste à sa place, alors qu’un clivage s’opère au niveau des jonctions adhérentes (constriction actine/myosine/N-cadhérine/αPKC [protéine kinase Cα]) pour libérer la cellule de son ancrage apical. Lors de cette scission cellulaire, les centrioles se déplacent vers le corps cellulaire, alors que les marqueurs ciliaires analysés restent dans la particule apicale. Finalement, cette perte rapide de polarité apicale permet au futur neurone de se déplacer dans l’épaisseur du neuroépithélium, d’acquérir de nouvelles caractéristiques morphologiques et fonctionnelles, bref de se différencier, et cela selon toute vraisemblance en l’absence de cil primaire, au moins dans un premier temps. Le devenir de la particule apicale n’a pas été exploré, mais elle pourrait être libérée dans la lumière du tube neural qui contient le liquide cérébrospinal (LCS), ce dernier véhiculant, de fait, toutes sortes de composants moléculaires et vésiculaires [
36]. Parmi ces composants, certains pourraient être produits par le cil lui-même, comme semblent l’indiquer les figures de bourgeonnement observées à l’extrémité du cil [
37]. Si cette fonction de sécrétion se confirmait, cela impliquerait que les cils ne sont pas seulement des antennes réceptrices, mais qu’ils pourraient également participer à la production de signaux.Protéine ciliaire ARL13B et polarité apico-basale Par ailleurs, l’établissement de la polarité apicobasale des cellules neuroépithéliales au début de la neurogenèse corticale dépend de la protéine ciliaire Arl13b (ADP-ribosylation factor-like 13B) (GTPase de la famille ARF/ARL) [
38]. Chez le mutant nul Arl13bhennin
, la polarité se trouve complètement inversée : le pôle apical se retrouve en périphérie et le pôle basal fait face aux cavités ventriculaires. Conformément à cette inversion, les progéniteurs sont en périphérie et les neurones corticaux migrent et se différencient successivement dans des couches plus internes, en parfaite opposition avec ce qui se passe en situation normale. L’utilisation de knock-out conditionnels a permis de montrer que Arl13b est critique pour l’acquisition de la polarité apico-basale entre E9,0 et E10,5. Néanmoins, il n’est pas certain que ce phénotype soit véritablement dû à un dysfonctionnement du cil. À ce jour, ce phénotype radical n’a pas été retrouvé chez d’autres mutants ciliaires (Ift88Cobblestone
, Rpgrip1l/Ftm, Kif3a, Rfx [regulatory factor 3X], etc.). En effet, l’identification d’un phénotype comme « ciliaire » réside dans son caractère commun, partagé avec d’autres mutants dont les anomalies ciliaires résultent de mutations différentes [65].Les différences phénotypiques, en revanche, peuvent être dues à des fonctions spécifiques de certaines protéines ciliaires, ou bien à des fonctions extraciliaires de la protéine mutée [ 70] (→). (→) Voir la Synthèse de N. Taulet et B. Delaval, page 1040 de ce numéro Or, nombre de fonctions non ciliaires (KIF3A, IFT88, etc.) ont été attribuées à des protéines ciliaires [ 39, 40]. En conclusion, la position du cil primaire semble refléter les propriétés de polarité apicobasale des cellules neurales et varie lors de leur différenciation. De plus, le cil, du fait de son lien intime avec le centrosome, semble également important dans l’organisation et la dynamique du cytosquelette, essentielles notamment aux migrations cellulaires des futurs neurones. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
Pendant longtemps, on a pensé que les cellules en migration étaient dépourvues de cil. On pensait que le centrosome ne pouvait pas s’ancrer à la membrane pour soutenir un cil alors qu’il était engagé dans l’organisation et le remaniement dynamique des microtubules conduisant à la progression des protrusions cellulaires et à la translocation nucléaire. Et pourtant, deux publications ont montré la présence et l’importance du cil primaire des interneurones pendant leur migration tangentielle vers le cortex [ 41, 42, 68] (→). (→) Voir la Synthèse de C. Métin, page 991 de ce numéro L’absence ou le dysfonctionnement du cil résultant de l’invalidation conditionnelle des gènes Kif3a, Ift88 ou Arl13b perturbent la migration des interneurones, en causant un allongement des périodes de pause, une augmentation du nombre de neurites et une stabilisation de ces protrusions. De plus, l’orientation de la migration est altérée dans le cortex, ce qui conduit à un positionnement anormal des interneurones dans les couches corticales. La voie de signalisation Shh semble impliquée dans ces défauts de migration, mais elle n’est probablement pas la seule en cause. Enfin, dès leur positionnement dans les différentes couches du cortex, les neurones pyramidaux vont produire des axones dont la croissance orientée permet l’établissement de connexions synaptiques avec d’autres neurones corticaux (projections cortico-corticales) ou avec des cibles se trouvant dans des territoires postérieurs du SNC (projections corticofuges) [ 43]. La grande majorité des projections cortico-corticales interhémisphériques transitent par le corps calleux (Figure 3) [ 44]. Or, des agénésies du corps calleux sont fréquemment observées dans certaines ciliopathies (OFD, syndrome de Jeune, syndrome de Meckel, syndrome acrocalleux) (Tableau I) [8]. L’analyse de deux mutants ciliaires, Rfx3 et Rpgrip1l/Ftm, a révélé une agénésie du corps calleux résultant de défauts de positionnement des cellules de guidage qui jalonnent la trajectoire empruntée par les axones calleux. Ces cellules se différencient en position ectopique chez les mutants ciliaires, probablement suite au défaut de régionalisation précoce du télencéphale. Ce phénotype, semblable à celui qui est décrit chez les mutants Gli3 [22], peut être corrigé par la réintroduction de Gli3-R chez les doubles mutants Ftm-/- Gli3∆699 et Rfx3-/- Gli3∆699 [ 45] (C. Laclef, manuscrit en préparation). De plus, chez ces mutants, existent également des défauts des projections corticofuges (structure très anormale du striatum), dont l’origine reste à analyser. Enfin, chez le mutant Bbs8, un défaut de fasciculation et de projection des axones issus de neurones sensoriels olfactifs a été mis en évidence [ 46]. Ainsi, de nombreux défauts de projections axonales sont en cours d’étude chez les modèles murins de ciliopathies, qui sont à mettre en parallèle avec les défauts de décussation (croisement de la ligne médiane), décrits notamment chez des patients atteints du syndrome de Joubert [ 47, 48, 65]. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
Une ventriculomégalie (dilatation des cavités ventriculaires) est associée à certaines ciliopathies (BBS, HLS [hydrolethalus syndrome] ; voir Tableau I ), mais son origine chez l’homme est probablement distincte de celle des hydrocéphalies identifiées chez les mutants ciliaires murins (Iftl88ORPK , Rpgrip1lFtm , Rfx3, Inpp5e, etc.). L’hydrocéphalie est une ventriculomégalie due à une augmentation de la pression intraventriculaire, qui peut avoir des conséquences dramatiques sur le tissu neural adjacent. On distingue les hydrocéphalies obstructives et communicantes, selon qu’il y a, ou non, occlusion des voies de circulation du liquide céphalospinal. Parmi les multiples causes d’hydrocéphalie postnatale citons les défauts des battements ciliaires coordonnés qui mettent en mouvement le liquide céphalospinal. Mais, chez la souris, ces cils mobiles qui tapissent les ventricules ne se forment que lorsque les cellules épendymaires se différencient, c’est-à-dire dans les deux semaines qui suivent la naissance [ 49]. Ils ne peuvent donc pas être en cause dans les hydrocéphalies anténatales. Dans ce cas, celles-ci peuvent s’expliquer par un déséquilibre entre la production (excessive) et la résorption (insuffisante) du liquide céphalospinal. Les cellules du plexus choroïde, qui sécrètent le liquide céphalospinal, portent des touffes de cils primaires, une caractéristique atypique, car les cils primaires sont en général solitaires [ 50]. Ces cils primaires réguleraient à la fois la quantité de liquide céphalospinal sécrété (par un mécanisme impliquant le récepteur au neuropeptide FF) et sa composition (accumulation d’ions chlorure chez les mutants Ift88ORPK ) [50, 51]. Par ailleurs, l’hydrocéphalie des souris Bbs1M390R/M390R , caractérisées par la mutation la plus fréquemment identifiée chez les patients BBS, peut être partiellement corrigée (50 %) par l’administration de lithium aux mères pendant la gestation [ 52]. Le lithium avait déjà été mentionné comme agoniste de la voie Wnt, capable de corriger des défauts de prolifération au niveau du cervelet, chez le mutant Ahi1 [ 53]. L’ion lithium est connu pour ses effets pléiotropes : il agit notamment sur la régulation de l’activité de la sérine thréonine kinase, GSK3β (glycogen synthase kinase 3β), impliquée, entre autres, dans les voies de signalisation PDGFα (plateled-derived growth factor α), Wnt/β-caténine et dans le clivage de Gli3 (voie Hh). Ces travaux ouvrent donc des perspectives thérapeutiques intéressantes, mais la compréhension des mécanismes sous-jacents nécessitera des études plus approfondies. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
Le cil primaire intervient clairement dans différentes étapes du développement cérébral anténatal, mais également dans le cerveau postnatal. La maturation des neurones se poursuit après la naissance, notamment avec l’établissement des connexions synaptiques. Or, il semble exister une corrélation entre la richesse des arborisations dendritiques et le cil primaire [ 54, 55]. Certaines régions cérébrales contiennent des cellules souches neurales dotées d’un cil qui génèrent de nouveaux neurones pendant toute la vie adulte. Or, chez les mutants Kif3a et Stumpy, la neurogenèse dans l’hippocampe est réduite, conséquence d’un défaut de signalisation Hh [ 56, 57]. Les cils primaires ont été décrits sur différents types cellulaires dans quasiment toutes les régions du cerveau adulte et ils sont souvent enrichis en récepteurs membranaires couplés aux protéines G [ 58, 59]. Indirectement, l’étude des mutants ciliaires obèses a permis de définir une fonction du cil dans le contrôle hypothalamique de la satiété [ 60, 61, 71] (→). (→) voir la Synthèse de K. Chennen et al., page 1034 de ce numéro Mais, de manière générale, le rôle joué par ce petit organite dans un océan de dendrites et d’axones reste encore bien mystérieux. Ce petit compartiment isolé pourrait-il concentrer et intégrer des signaux annexes, extrasynaptiques, qui contribuent à la survie, à la plasticité, voire à la fonction des neurones ? Plusieurs maladies neurodégénératives ont été associées à des dysfonctionnements ciliaires. La dégénérescence des photorécepteurs de la rétine est très fréquente dans les ciliopathies. Le cil pourrait aussi être en cause dans les ataxies spinocérébelleuses [26, 62], la chorée de Huntington, la maladie de Parkinson, ou encore certains syndromes neurologiques tels que la schizophrénie ou la dyslexie [2, 5]. Les recherches futures, notamment l’exploitation d’animaux chez lesquels un knock-out conditionnel permettrait de cibler l’invalidation des gènes ciliaires dans le cerveau adulte, devraient offrir un nouvel éclairage sur ces questions. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
L’auteur déclare n’avoir aucun lien d’intérêt concernant les données publiées dans cet article. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
Je remercie les membres du laboratoire pour leur relecture attentive du manuscrit et les critiques constructives qu’ils m’ont faites : Sylvie Schneider-Maunoury, Christine Vesques, Guillaume Pezeron et bien sûr Martin Catala pour la partie médicale. Merci aussi à Christine Métin pour ses commentaires avisés. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||



