 |
| |||
Med Sci (Paris). 2014 April; 30(4): 398–404. Published online 2014 May 5. doi: 10.1051/medsci/20143004013.Tissu adipeux et cancer Une association à haut risque 1Institut de pharmacologie et de biologie structurale, CNRS/université de Toulouse, UMR 5089, 205, route de Narbonne, BP 64182, 31077Toulouse, France 2Inserm U1048, Institut des maladies métaboliques et cardiovasculaires de Toulouse (I2MC), 1, avenue Jean Poulhès, BP 84225, 31432Toulouse Cedex 4, France Corresponding author. | ||||||
| ||||||
Le cancer doit être maintenant considéré comme une maladie tissulaire au cours de laquelle les cellules tumorales interagissent de façon dynamique avec leur micro-environnement ou stroma [ 1]. Cet environnement est principalement composé de fibroblastes, de cellules immunes et endothéliales, et d’une matrice extracellulaire (MEC) (→) [ 37, 38]. Au cours de la progression tumorale, les cellules cancéreuses vont profondément modifier leur microenvironnement qui, en retour, va favoriser la croissance et la dissémination de la tumeur. L’importance dans ce processus des fibroblastes associés au cancer (CAF pour cancer-associated fibroblast) ou encore des macrophages (TAM pour tumor-associated macrophage) a été clairement décrite [ 2, 3]. (→) Voir les Synthèses de S. Le Guellec et al., et de S. Provot, pages 366 et 372 de ce numéro Les cellules qui composent le tissu adipeu sont aussi fréquemment retrouvées à proximité des tumeurs, en particulier dans le cancer du sein où elles sont un des composants majoritaires du stroma tumoral. Leur rôle a été longtemps ignoré, mais l’augmentation du nombre d’études les concernant souligne l’intérêt croissant de la communauté scientifique pour cette nouvelle facette du micro-environnement tumoral. Pourquoi s’intéresser au tissu adipeux et, plus particulièrement, aux adipocytes matures ? Ces dernières années, il a été montré que les adipocytes n’étaient pas de simples réservoirs d’énergie, mais des cellules endocrines capables de sécréter une grande variété de molécules (appelées adipokines), incluant des hormones, des facteurs de croissance, des chimiokines ou des molécules pro-inflammatoires, sécrétions dont le profil est modifié dans des conditions d’obésité [ 4]. Ce profil sécrétoire fait donc des adipocytes d’excellents candidats pour influencer le comportement des tumeurs. Le lien clairement établi entre obésité et cancer renforce l’intérêt d’étudier le rôle des adipocytes matures dans la progression tumorale. En effet, des études épidémiologiques ont maintenant montré de façon convaincante que l’obésité est associée à une augmentation de la mortalité par cancer [ 5]. Ces données ont des conséquences majeures en termes de santé publique, puisque le surpoids et l’obésité sont en augmentation constante dans le monde, et que leur prévalence atteint maintenant des dimensions alarmantes. En effet, l’Organisation mondiale de la santé prévoit qu’en 2015, trois milliards d’adultes dans le monde seront en surpoids, et 700 millions seront obèses. Le lien entre obésité et mortalité par cancer s’explique à la fois par une augmentation de leur incidence (en particulier celle des cancers de l’endomètre, de l’œsophage, du côlon, du rein et du sein en période post-ménopausique), de leur agressivité (cancer du sein et de la prostate principalement) chez les sujets obèses [5]. Dans le cas du cancer du sein et indépendamment du statut ménopausique, l’obésité est un facteur indépendant de mauvais pronostic [ 6], les femmes obèses présentant des tumeurs à un stade avancé et qui métastasent plus fréquemment (pour revue [ 7, 8]). Ce dialogue délétère entre les cellules tumorales et les adipocytes pourrait donc être amplifié au cours de l’obésité où l’équilibre normal des protéines sécrétées par le tissu adipeux est perturbé. | ||||||
Le tissu adipeux constitue le lieu privilégié de mise en réserve des lipides. On regroupe sous ce terme deux tissus très distincts : le tissu adipeux brun et le tissu adipeux blanc [ 36]. Le tissu adipeux brun, dont la présence chez l’homme a été décrite depuis un certain nombre d’années, génère un regain d’intérêt depuis l’apparition de technologies, comme la tomographie par émission de positons (TEP), qui ont permis de révéler, via son activité métabolique, la persistance de ce tissu chez l’homme adulte [ 39]. Ce tissu adipeux a pour rôle majeur de libérer de l’énergie sous forme de chaleur [ 9]. Ce tissu adipeux n’ayant pas été pour l’instant impliqué dans le dialogue entre adipocytes et cellules cancéreuses, il n’en sera pas question dans cette revue. Cependant, il est intéressant de noter que le tissu adipeux brun semble impliqué dans la cachexie cancéreuse [ 10]. Le rôle essentiel du tissu adipeux blanc est d’assurer le maintien de l’équilibre énergétique en stockant l’énergie sous forme de triglycérides ou en la libérant sous forme d’acides gras. Un adipocyte blanc, caractérisé par son aspect sphérique, contient une large et unique vacuole lipidique (composée à 95 % de triglycérides), dont la taille est susceptible d’augmenter ou de diminuer assez rapidement en fonction de la balance énergétique. Outre les adipocytes matures, on retrouve, dans une fraction dite stroma vasculaire du tissu adipeux, de nombreuses cellules, telles que des cellules progénitrices (adipose derived stem cells ou ADSC et pré-adipocytes), des fibroblastes, des macrophages, des lymphocytes, des péricytes et des cellules endothéliales. La MEC permet la cohésion du tissu adipeux et joue un rôle important dans la régulation du développement et du métabolisme des adipocytes [ 11]. L’expression « le tissu adipeux » n’a pas vraiment de sens, car ce tissu n’est pas homogène et présente des spécificités anatomiques et fonctionnelles, selon sa localisation et l’importance de son extension. Chez les mammifères, le tissu adipeux blanc est disséminé dans des sites anatomiques divers. Les localisations sous-cutanées (fémoro-glutéale et abdominale), intra-abdominales/périviscérales (omentale, rétro-péritonéale) et périgonadiques (épididymaire, péri-ovarienne) sont les plus importantes. On trouve enfin des adipocytes dans la glande mammaire (où ils fournissent l’énergie nécessaire à la lactation) et dans la moelle osseuse [11] (Figure 1). Le métabolisme adipocytaire varie selon le site anatomique considéré et peut ainsi conditionner l’augmentation de certains dépôts adipeux. Par exemple, le tissu adipeux périviscéral est bien plus réactif à la mobilisation lipidique que le tissu adipeux sous-cutané [ 12]. Outre sa fonction métabolique, comme nous l’avons vu précédemment, l’adipocyte possède un formidable potentiel sécrétoire. Il s’agit, outre les acides gras libres (AGL), produit de sécrétion majoritaire, de très nombreuses protéines (Figure 2) [4]. À la leptine, qui a donné ses lettres de noblesse au tissu adipeux par son activité sur le système nerveux central (action sur le contrôle de la prise alimentaire et la thermorégulation), se sont rapidement ajoutées d’autres molécules impliquées dans le métabolisme énergétique, comme l’adiponectine ou, plus récemment, l’apeline [ 13]. Les adipocytes sécrètent aussi d’autres hormones (stéroïdes, glucocorticoïdes), certains lipides (prostaglandines, acide lysophosphatidique), des molécules pro-angiogéniques (monobutyrine, VEGF [vascular endothelial growth factor], etc.), des chimiokines (comme MCP1, pour monocyte-chemoattracting protein 1) et des facteurs de croissance (tels que l’HGF pour hepatocyte growth factor). De nombreuses cytokines pro-inflammatoires, comme le TNFα (tumor necrosis factor α) ou certaines interleukines (IL1β, IL6) sont également produites par la cellule adipeuse. Le profil sécrétoire des adipocytes varie en fonction de la localisation du tissu adipeux. La leptine, mais aussi l’adiponectine, sont principalement sécrétées par le tissu adipeux sous-cutané, alors que le tissu adipeux périviscéral est la principale source de cytokines pro-inflammatoires et de peptides bioactifs, comme le VEGF (pour revue [ 14]). Les conséquences de l’expansion des différents dépôts adipeux sont aussi différentes. Ainsi, l’augmentation du tissu adipeux périviscéral est particulièrement liée aux complications de l’obésité, telles que le diabète de type II, les maladies cardiovasculaires et les dyslipidémies, alors que l’augmentation du tissu adipeux sous-cutané pourrait s’avérer protectrice vis-à-vis de ces dernières. Il est maintenant clairement établi que l’obésité est associée à un état inflammatoire à bas bruit dû à une infiltration de macrophages dans le tissu adipeux périviscéral, et que cet état joue un rôle majeur dans les complications de cette maladie [ 15].
Les spécificités fonctionnelles des différents tissus adipeux soulignent l’importance de la nature du tissu adipeux utilisé dans les études expérimentales analysant le dialogue entre adipocytes et cellules cancéreuses. La plupart des études réalisées à ce jour l’ont été avec des lignées pré-adipocytaires murines (3T3-L1 ou 3T3-F442A) qui se différencient, à confluence, en adipocytes fonctionnels. Ces lignées ont l’avantage d’être faciles d’utilisation et de reproduire assez fidèlement l’interaction entre cellules cancéreuses et adipocytes, comme nous l’avons montré dans le cancer du sein [ 16]. Toutefois, il convient de vérifier les effets observés avec des adipocytes obtenus à partir du tissu adipeux d’intérêt. Il peut s’agir d’adipocytes matures différenciés ex vivo à partir de progéniteurs adipocytaires, ou isolés et maintenus in vitro dans des systèmes de coculture en 3D ; la durée de vie de ces cellules en 2D est en effet extrêmement limitée [8, 17]. Enfin, la validation des modifications adipocytaires observées se doit d’être effectuée dans les tumeurs humaines. | ||||||
Rôle paracrine des adipocytes dans la progression tumorale : le concept de cancer-associated adipocytes
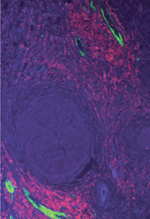
Le rôle du microenvironnement dans la progression tumorale s’explique largement par la mise en place d’un dialogue bidirectionnel entre les cellules cancéreuses et les cellules « normales » environnantes qui peuvent être activées localement ou recrutées à distance. C’est dans le cancer du sein que la liaison dangereuse qui s’établit entre tumeurs et adipocytes a été le mieux caractérisée. C’est dans les années 1990, que le rôle du tissu adipeux dans la croissance tumorale et la formation de métastases a été évoqué [ 18], mais il a fallu plus de 10 ans pour le démontrer [ 19]. Aujourd’hui, les molécules impliquées dans le dialogue entre adipocytes et cancer commencent à être bien identifiées : cytokines pro-inflammatoires, protéines de la MEC ou de son remodelage, AGL [8, 20, 21]. Afin de modéliser l’interaction entre adipocytes et cancer, nous avons mis en place au laboratoire un modèle original de coculture en 2D où les deux populations sont séparées par un insert autorisant la diffusion de facteurs solubles [16]. Dans ces conditions, l’invasion des cellules tumorales est fortement stimulée, qu’il s’agisse de lignées humaines ou murines exprimant ou non le récepteur aux œstrogènes (RE), alors que l’effet sur la prolifération semble plus dépendant du modèle cellulaire utilisé. Les adipocytes cocultivés avec des cellules tumorales présentent un phénotype spécifique caractérisé à la fois par : (1) une délipidation et une diminution de l’expression des marqueurs adipocytaires, (2) la surexpression de molécules pro-inflammatoires comme l’IL6, et (3) la surexpression de protéines de la MEC ou impliquées dans son remodelage comme la MMP11. Par analogie aux CAF et aux TAM, nous avons nommé ces cellules CAA (cancer-associated adipocyte) [7] (Figure 3). Ainsi, il existe bien un adipocyte péritumoral, dont le phénotype et les sécrétions sont différents de ceux d’un adipocyte « naïf ». Au front invasif des tumeurs mammaires, on retrouve des adipocytes de petite taille correspondant à des CAA (Figure 4B) et dont le profil de sécrétion est similaire à ce qui a été décrit in vitro dans le modèle de coculture, validant ainsi l’intérêt de ce modèle [16, 22]. Ces modifications de la taille et du contenu lipidique des adipocytes sont observées au niveau de la zone de contact entre tissu adipeux et la plupart des tumeurs invasives, comme l’illustre la Figure 4 pour les cancers de la prostate et les mélanomes (données de notre équipe) ou le cancer de l’ovaire avec dissémination péritonéale [ 23].
Trois grands types de modifications caractérisent donc les CAA.
Un autre aspect original et très intéressant de ce dialogue entre adipocytes et cancer est l’apparition d’une résistance aux traitements. Nous avons récemment démontré que la coculture de lignées tumorales mammaires en présence d’adipocytes favorisait l’apparition d’un phénotype de radiorésistance [ 30]. D’autre part, il a également été montré in vitro et in vivo que les adipocytes promouvaient la chimiorésistance dans un modèle de leucémie lymphoblastique [ 31]. Enfin, l’inhibition de l’endotrophine, le produit de clivage du Col6, sensibilise les tumeurs au cisplatine [ 32]. Le dialogue entre adipocytes et cancer fait aussi intervenir d’autres adipokines, telles que la leptine et l’adiponectine, qui n’ont pas été traitées dans cette revue. Ces adipokines pourraient agir à un niveau paracrine mais certainement aussi endocrine, comme la plupart des sécrétions adipocytaires. Le lecteur intéressé par cet aspect de l’interaction entre tissu adipeux et cancer peut se référer à la revue de Park et al. [ 33]. De plus, bien que l’effet pro-invasif des adipocytes péritumoraux soit indépendant de l’expression du récepteur des œstrogènes, la production d’œstrogènes par les adipocytes (via leur capacité d’aromatisation des androgènes) est susceptible de favoriser la prolifération de certaines tumeurs RE+ et participe certainement à l’initiation tumorale dans le cas des cancers du sein survenant après la ménopause [ 34]. Enfin, d’autres composants du tissu adipeux, et en particulier les ADSC, contribuent à la progression tumorale, ce qui renforce l’importance de l’ensemble des composants de ce tissu en cancérologie [8]. | ||||||
Comme nous l’avons vu dans le premier paragraphe, l’obésité est considérée comme un facteur de mauvais pronostic indépendant, en particulier dans le cancer du sein [5]. Nous proposons l’hypothèse selon laquelle les adipocytes présents à proximité des tumeurs seraient, chez l’obèse, encore plus enclins à stimuler la dissémination locale et à distance, ce qui explique le pronostic défavorable observé chez ces patientes [7, 8, 16]. Il est aujourd’hui admis que le tissu adipeux, chez l’obèse, est dans un état d’inflammation dit à bas bruit, participant aux nombreux effets délétères de l’obésité. La présence de zones de nécrose adipocytaire (favorisant la libération d’AGL) entourées de macrophages (crown-like structure ou CLS) et de taux élevés de cytokines pro-inflammatoires sont des marqueurs couramment utilisés pour mettre en évidence cet état sub-inflammatoire du tissu adipeux périviscéral chez le sujet obèse [11]. De façon très intéressante, des études récentes ont montré, dans des modèles murins et chez l’homme, que le tissu adipeux mammaire des sujets obèses se caractérise par une hypertrophie adipocytaire, des CLS (crown-like structures) et une augmentation de la sécrétion de cytokines inflammatoires (pour revue [8]). Outre cet état sub-inflammatoire, d’autres modifications fonctionnelles sont observées dans le tissu adipeux des sujets obèses, telles que l’hypoxie et l’apparition de zones de fibrose [ 35] qui sont elles aussi susceptibles d’influencer l’interaction avec la tumeur [40] (→). Il est donc intéressant ici de souligner la similitude qui existe entre les caractéristiques des CAA (sécrétion de cytokines pro-inflammatoires, augmentation de la sécrétion de protéines matricielles, libération d’AGL) et celle des adipocytes dans un contexte d’obésité, ce qui renforce notre hypothèse. Il existe à l’heure actuelle toutefois peu d’arguments expérimentaux. Les modèles animaux montrent de façon convaincante que l’obésité (induite par un régime lipidique) s’accompagne d’une augmentation de la progression tumorale dans différents modèles, sans toutefois que le caractère paracrine ou endocrine de cet effet n’ait été démontré (pour revue [21]). La caractérisation tissulaire des tumeurs chez les sujets obèses et dans des modèles animaux d’obésité, associée à une modélisation de cette interaction dans des modèles de coculture en 3D utilisant des adipocytes isolés de tissu adipeux d’obèses, devrait permettre de valider ou non cette hypothèse. (→) Voir la Synthèse de M. Hasmim et al., page 422 de ce numéro En conclusion, les travaux de ces dix dernières années ont permis d’établir que les adipocytes sont un composant important du microenvironnement tumoral, ce qui pourrait permettre de mieux appréhender à terme les liens entre nutrition, obésité et cancer. L’évolution du surpoids et de l’obésité, et donc l’augmentation attendue dans un proche avenir de l’incidence de cancers agressifs liés à l’obésité ne font que souligner l’importance des recherches dans ce domaine afin d’élaborer des stratégies originales visant à prévenir et traiter ces cancers chez ce sous-groupe spécifique de patients. | ||||||
Les auteurs déclarent n’avoir aucun lien d’intérêt concernant les données publiées dans cet article. | ||||||
Les travaux réalisés dans nos équipes ont bénéficié du soutien financier de l’INCA (INCA PL 2006–035 and INCA PL 2010-214, PV et CM), de la Ligue régionale Midi-Pyrénées contre le cancer (comité du Lot, de la Haute-Garonne et du Gers à CM), de la Fondation de France (PV et CM), de l’Association pour la recherche sur les tumeurs prostatiques ARTP (CM) et de l’université de Toulouse (appel d’offres du conseil scientifique 2009 à CM). Victor Laurent bénéficie d’une bourse de thèse de l’ARC. Nous remercions Françoise Viala pour son aide dans la réalisation de l’iconographie de ce manuscrit. | ||||||
1.
Allen
M
,
Louise Jones
J
. Jekyll and Hyde: the role of the microenvironment on the progression of cancer . J Pathol.
2011; ; 223 : :162.–176. 2.
Polanska
UM
,
Orimo
A
. Carcinoma-associated fibroblasts: non-neoplastic tumour-promoting mesenchymal cells . J Cell Physiol.
2013; ; 228 : :1651.–1657. 3.
Ruffell
B
,
Affara
NI
,
Coussens
LM
. Differential macrophage programming in the tumor microenvironment . Trends Immunol.
2012; ; 33 : :119.–126. 4.
Rajala
MW
,
Scherer
PE
. Minireview: The adipocyte-at the crossroads of energy homeostasis, inflammation, and atherosclerosis . Endocrinology.
2003; ; 144 : :3765.–3773. 5.
Calle
EE
,
Kaaks
R
. Overweight, obesity and cancer: epidemiological evidence and proposed mechanisms . Nat Rev Cancer.
2004; ; 4 : :579.–591. 6.
Parekh
N
,
Chandran
U
,
Bandera
EV
. Obesity in cancer survival . Annu Rev Nutr.
2012; ; 32 : :311.–342. 7.
Dirat
B
,
Bochet
L
,
Escourrou
G
, et al.
Unraveling the obesity and breast cancer links: a role for cancer-associated adipocytes?
Endocr Dev.
2010; ; 19 : :45.–52. 8.
Wang
YY
,
Lehuede
C
,
Laurent
V
, et al.
Adipose tissue and breast epithelial cells: a dangerous dynamic duo in breast cancer . Cancer Lett.
2012; ; 324 : :142.–151. 9.
Smorlesi
A
,
Frontini
A
,
Giordano
A
,
Cinti
S.
, The adipose organ: white-brown adipocyte plasticity, metabolic inflammation . Obes Rev.
2012; ; 13 : (suppl 2) : :83.–96. 10.
Tsoli
M
,
Moore
M
,
Burg
D
, et al.
Activation of thermogenesis in brown adipose tissue and dysregulated lipid metabolism associated with cancer cachexia in mice . Cancer Res.
2012; ; 72 : :4372.–4382. 11.
Ouchi
N
,
Parker
JL
,
Lugus
JJ
,
Walsh
K
. Adipokines in inflammation and metabolic disease . Nature Rev Immunol.
2011; ; 11 : :85.–97. 12.
Lafontan
M
,
Langin
D
. Lipolysis and lipid mobilization in human adipose tissue . Prog Lipid Res.
2009; ; 48 : :275.–297. 13.
Boucher
J
,
Masri
B
,
Daviaud
D
, et al.
Apelin, a newly identified adipokine up-regulated by insulin and obesity . Endocrinology.
2005; ; 146 : :1764.–1771. 14.
Galic
S
,
Oakhill
JS
,
Steinberg
GR
. Adipose tissue as an endocrine organ . Mol Cell Endocrinol.
2010; ; 316 : :129.–139. 15.
Fain
JN
. Release of interleukins and other inflammatory cytokines by human adipose tissue is enhanced in obesity and primarily due to the nonfat cells . Vitam Horm.
2006; ; 74 : :443.–477. 16.
Dirat
B
,
Bochet
L
,
Dabek
M
, et al.
Cancer-associated adipocytes exhibit an activated phenotype and contribute to breast cancer invasion . Cancer Res.
2011; ; 71 : :2455.–2465. 17.
Lafontan
M
. Historical perspectives in fat cell biology: the fat cell as a model for the investigation of hormonal and metabolic pathways . Am J Physiol Cell Physiol.
2012; ; 302 : :C327.–C359. 18.
Elliott
BE
,
Tam
SP
,
Dexter
D
,
Chen
ZQ
. Capacity of adipose tissue to promote growth and metastasis of a murine mammary carcinoma: effect of estrogen and progesterone . Int J Cancer.
1992; ; 51 : :416.–424. 19.
Manabe
Y
,
Toda
S
,
Miyazaki
K
,
Sugihara
H
. Mature adipocytes, but not preadipocytes, promote the growth of breast carcinoma cells in collagen gel matrix culture through cancer-stromal cell interactions . J Pathol.
2003; ; 201 : :221.–228. 20.
Hefetz-Sela
S
,
Scherer
PE
. Adipocytes: Impact on tumor growth and potential sites for therapeutic intervention . Pharmacol Ther.
2013; ; 138 : :197.–210. 21.
Nieman
KM
,
Romero
IL
,
Van Houten
B
,
Lengyel
E
. Adipose tissue and adipocytes support tumorigenesis and metastasis . Biochim Biophys Acta.
2013; ; 1831 : :1533.–1541. 22.
Andarawewa
KL
,
Motrescu
ER
,
Chenard
MP
, et al.
Stromelysin-3 is a potent negative regulator of adipogenesis participating to cancer cell-adipocyte interaction/crosstalk at the tumor invasive front . Cancer Res.
2005; ; 65 : :10862.–10871. 23.
Nieman
KM
,
Kenny
HA
,
Penicka
CV
, et al.
Adipocytes promote ovarian cancer metastasis and provide energy for rapid tumor growth . Nat Med.
2011; ; 17 : :1498.–1503. 24.
Finley
DS
,
Calvert
VS
,
Inokuchi
J
, et al.
Periprostatic adipose tissue as a modulator of prostate cancer aggressiveness . J Urol.
2009; ; 182 : :1621.–1627. 25.
Wagner
M
,
Bjerkvig
R
,
Wiig
H
, et al.
Inflamed tumor-associated adipose tissue is a depot for macrophages that stimulate tumor growth and angiogenesis . Angiogenesis.
2012; ; 15 : :481.–495. 26.
Iyengar
P
,
Espina
V
,
Williams
TW
, et al.
Adipocyte-derived collagen VI affects early mammary tumor progression in vivo, demonstrating a critical interaction in the tumor/stroma microenvironment . J Clin Invest.
2005; ; 115 : :1163.–1176. 27.
Park
J
,
Scherer
PE
. Adipocyte-derived endotrophin promotes malignant tumor progression . J Clin Invest.
2012; ; 122 : :4243.–4256. 28.
Motrescu
ER
,
Blaise
S
,
Etique
N
, et al.
Matrix metalloproteinase-11/stromelysin-3 exhibits collagenolytic function against collagen VI under normal and malignant conditions . Oncogene.
2008; ; 27 : :6347.–6355. 29.
Gazi
E
,
Gardner
P
,
Lockyer
NP
, et al.
Direct evidence of lipid translocation between adipocytes and prostate cancer cells with imaging FTIR microspectroscopy . J Lipid Res.
2007; ; 48 : :1846.–1856. 30.
Bochet
L
,
Meulle
A
,
Imbert
S
, et al.
Cancer-associated adipocytes promotes breast tumor radioresistance . Biochem Biophys Res Commun.
2011; ; 411 : :102.–106. 31.
Behan
JW
,
Yun
JP
,
Proektor
MP
, et al.
Adipocytes impair leukemia treatment in mice . Cancer Res.
2009; ; 69 : :7867.–7874. 32.
Park
J
,
Morley
TS
,
Scherer
PE
. Inhibition of endotrophin, a cleavage product of collagen VI, confers cisplatin sensitivity to tumours . EMBO Mol Med.
2013; ; 5 : :935.–948. 33.
Park
J
,
Euhus
DM
,
Scherer
PE
. Paracrine and endocrine effects of adipose tissue on cancer development and progression . Endocrine Rev.
2011; ; 32 : :550.–570. 34.
Rose
DP
,
Vona-Davis
L
. The cellular and molecular mechanisms by which insulin influences breast cancer risk and progression . Endocrine Relat Cancer.
2012; ; 19 : :R225.–R241. 35.
Divoux
A
,
Clement
K
. Architecture and the extracellular matrix: the still unappreciated components of the adipose tissue . Obesity Rev.
2011; ; 12 : :e494.–e503. 36.
Carrière
A
,
Jeanson
Y
,
Cousin
B
, et al.
Le recrutement et l’activation d’adipocytes bruns et/ou BRITE : une perspective réelle pour le traitement des maladies métaboliques ?
Med Sci (Paris).
2013; ; 29 : :729.–735. 37.
Le Guellec
S
,
Duprez-Paumier
R
,
Lacroix-Triki
M
. Microenvironnement tumoral : la vision du pathologiste . Med Sci (Paris).
2014; ; 30 : :372.–377. | ||||||




